Ethan Linck
Cataclysms to Desolate the World
ISSUE 96 | PROPHECIES | JAN 2021

In Our Bodies, In Our Families by Ravi Zupa.
Though I knew they were coming, the sheer volume of spiderwebs still took me by surprise, stretching from the boughs of low trees to knuckles of sharp karst only partly buried in leaf litter. On Guam, the forests have no birds, and nature abhors a vacuum.
Even for the tropical Pacific—a region rent by successive waves of extinction that followed human expansion like the wake of an aircraft carrier—Guam’s missing birds and surplus spiderwebs are signs of an unusual legacy of loss. In the span of about 20 years, from the mid-1960s to 1985, a handful of brown tree snakes that had originally hitched a ride to the island from New Guinea during World War II exploded in number and managed to exterminate the entire native forest bird community. As the forests emptied and the branches of trees with names like ifit and nunu no longer served as perches for flycatchers and kingfishers and fantails, insect populations exploded, yearning for something to eat them. Soon after came the spiders.
The story is rich with symbolism, which is probably why it has become a textbook example of extinction. The West, by accident of imperial war, introduces a snake into an edenic Pacific paradise, with catastrophic consequences. Birds, whose beauty and ubiquity have always given them a special status among animals, disappear at the hand of our folly, and are replaced with a primal horror. The balance of nature, once disturbed, is difficult to regain.
The moral of the fable is clear—yet its tidiness is deceptive, even dangerous. Who exactly is at fault? What balance was disrupted? Biology is a famously imprecise science, where variation is the only rule and cause and effect can be hopelessly confounded. At the same time, its models and theories seem especially prone to simplistic, deterministic misinterpretations—for example, the very idea that a given species’ extinction is an inevitable consequence of its fitness, that a species’ evolutionary past could prophesy its future.
I had come to Guam during the summer before my junior year of college, a research assistant for a team focused on the bigger picture: what happened to a forest where fruit doves no longer ate fruit and dispersed seeds, a forest no longer filled with their mournful, cascading calls? As we identified plants in the understory and measured the diameter of saplings, the silence would sit uneasily on me, and my mind would drift to a poster in the stairwell of the house our field team rented. It was a poster depicting the birds of the island, both those that had been saved by last minute interventions to establish captive populations—like the ko’ko’, or Guam rail—and those that had not, like the Guam flycatcher, or chuguangguang.
With slate-blue wings and white breast feathers fringed by delicate rust staining, the chuguangguang possessed a sort of a preemptively nostalgic beauty, and of all the species on the poster, it was the one that stuck in my head. For millions of years this small blue bird had flitted and breathed and caught mosquitoes and laid small pale eggs. And then one morning in 1984, there were no more. The tree of life had grown a little sparser, and standing there on the ragged limestone, in the heat and spiders and quiet, I wondered if it was always going to be this way.
* * *
Trees that depict evolutionary relationships among groups of organisms are called phylogenies. As abstractions, they are both visualizations and statistical objects. They help us make sense of complex patterns of kinship, and they convey unexpectedly complex arrays of quantitative information about the timing and frequency of events in the distant past. Importantly, they are hypotheses: they represent our informed hunches about a process we cannot observe directly, generated using computers and mathematical models that fit patterns we can detect: why we share similar bone structure to other primates, for example, or why both have highly similar stretches of DNA sequence.
Phylogenies are typically drawn as trees that spread from left to right or bottom to top, but we read them in the opposite direction: from the leaves (species alive today, or extinct species with no extant descendants) to the trunk (the common ancestor of all depicted lifeforms). Beginning with two species that are each other's closest relatives and moving left—conceptually, this is equivalent to moving back in time—their individual branches will eventually merge and become one. This junction represents a speciation event: the evolutionary process that split the common ancestor of these two species into the unique entities they are today.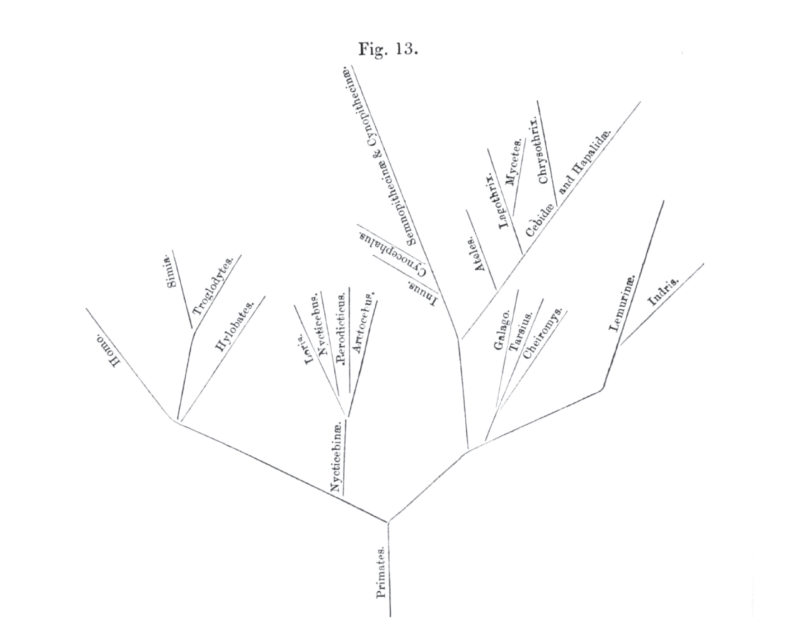
Phylogeny of primates, St. George Mivart 1865
Written this way, the interpretation of each tree seems unambiguous and intuitive. Yet even for experts, “treethinking” can be full of pitfalls. In spite of bumper-sticker misconception, there is no structured progression of life from “primitive” to “advanced” forms; evolution is frequently random and rarely consistently unidirectional. As representations of one aspect of the evolutionary process, phylogenies therefore contain no hierarchy of biological sophistication, though their very structure seems designed to trick us into believing there is, the patterns in their branches suggesting ladders, canopies, evolutionary staircases.
But there is another, more pervasive source of bias in what trees show us—and what they hide. The models used to build phylogenies reflect an idealized version of evolution, and the relationships in the trees they generate are uncertain. They are solutions for a puzzle we pose in the numerical universe of the model, a universe that may not share much in common with our own, and they may be upended when new data is discovered. Because we build phylogenies from things we can observe and count directly, they are necessarily an incomplete representation of the richness of biodiversity through space and time. If we’re very lucky, we can use evidence from the fossil remains of extinct plants and animals to generate hypotheses of where they fit into the bigger picture; for the vast majority of species that have left no trace in the fossil record, however, this is obviously impossible.
Those missing species, though, might easily slot into a phylogeny in unexpected places, changing our understanding of evolutionary relationships. And if extinction were pervasive enough, you could speculate, the placement of extinct species—and their frequency—might fundamentally alter the shape of a tree itself. Extinction, then, would leave a signature. The ghosts of the departed would linger among the living, not quite invisible.
It’s a line of speculation that poses an intriguing question: what can we learn from phylogenies by reading between the branches?
* * *
The chuguangguang was a monarch flycatcher, a member of a family of 100 or so small, insect-eating birds distributed from the western Pacific to Africa. Because of its diversity, the tree of monarch flycatchers is dense and bushy, foliated by species with names like Terpsiphone mutata and Arses telescopthalmus. The closest relative of the chuguangguang is the dark-grey flycatcher, Myiagra galeata, which lives on in the Moluccas, a chain of islands in western Indonesia some 1,500 miles away. They split from their common ancestor a million or so years ago, which itself had split from the common ancestor of a handful of other species another million or so years before.
If you were to count the total number of these splits throughout the tree, back until some arbitrary point in the past, and divide that by the number of years you covered in your bookkeeping, you could learn something useful: the speciation rate for that time period, or how quickly new species have been generated. If you were then to repeat this process for a similarly sized interval of time deeper in the tree, and then again and again until you reached its roots, you would get many such estimates of speciation rates. By comparing these rates, you could then learn about how the rate at which new species are generated through time has changed.
Recall, though, that these estimates will only be as good as the trees we use to make them. A time period where many extinct and undiscovered species originally diverged from their relatives will therefore show a lower speciation rate than actually occurred. For this reason, a better term for what you’re calculating is diversification rate—the net gain in new species after accounting for any losses. With sufficient statistical power, this dip in diversification rate will hint that at this point in the past, something interesting occurred. If we have reason to assume that the true rate of speciation has been constant through time, then we can infer that we are seeing the fingerprints of extinction.
I want to pause for a moment and reflect on this point. In the right combination of circumstances, the absence of evidence in a phylogeny can reveal itself as evidence of absence. These circumstances may or may not occur at a useful frequency—studying extinction with phylogenies is a perennially controversial topic, because many scientists are wary of the underlying assumptions—but in principle, it remains possible. Extinction leaves an ellipsis in the story of evolution. If, like countless other species throughout existence, the chuguangguang and a handful of its relatives had disappeared without ever being glimpsed by human eyes, we might still have guessed that there had been birds like them, once. We might still have wondered how they had lived, how they had shaped and been shaped by their small island world.
Explicitly considering extinction as a force shaping phylogenies also complicates a common epistemology of evolutionary thought. Tree thinking may not be hierarchical, but it is unavoidably linear and deterministic in its rendering of relationships as a bifurcating chain of ancestors and descendants. Extinction takes this chain and, by removing links, warps it. Malthus and the birth of industrial capitalism cast a long shadow in evolutionary biology—from Darwin’s early thinking on overpopulation to modern theories of “fitness landscapes,” the field has been dominated by ideas of competition, of struggle, and of the pursuit of some golden, optimal form. In contrast, acknowledging the interplay between extinction and speciation in phylogenies reflects a more dialectical mode of thought. It is the tension between the birth and death of species that has made the tree of life what we recognize it as today.
* * *
It’s hard to say when the fate of Guam’s birds was sealed, but there’s an argument to make that it was on March 6th, 1521, when the island was first spotted by Ferdinand Magellan. Though he had a dim view of the Marianas (which he called the “Islas de los Ladrones”) and the Indigenous Chamorro (“poor, but ingenious and very thievish”), they were nonetheless annexed by the Spanish Empire 44 years later.
The annexation of Guam and the rest of the Marianas began a history of violence and colonial exploitation that continues to this day. After four centuries under Spanish rule, the island was transferred to US control in 1898 following the Spanish-American War. It became a Navy base and a crucial link in the shipping network that drove the wheels of international capitalism. In World War II, Guam was invaded by the Japanese, who occupied it for 31 months. Allied Forces fought desperately to recapture it, and in the process almost completely stripped the island of trees with napalm. They were mostly replaced by tangan-tangan, a legume from South America. Haole biologists like me came from the metropolis to the periphery to study the ecological fallout of imperialism. And at some point, a handful of brown tree snakes slithered out from a shipping container, or uncurled themselves from a tank turret, and slipped into the jungle.
Of course, extinction is not only a symptom of modernity, and Guam’s avian losses in the 20th century were not its first. In 2009, paleontologists Gregory Pregill and David Steadman published a paper in the journal Diversity and Distributions with fossil evidence for five species previously unknown from the island—birds that had vanished prior to Magellan’s arrival. Curiously, two of these long-ago extinctions had been rails, like the ko’ko’—small, flightless birds that are more often heard than seen.
Rails seem to be particularly vulnerable to extinction and account for the majority of the 2,000 or so species of birds that have gone extinct in the past 20,000 years. Because of their basic biology, it’s easy to propose mechanisms for this vulnerability: maybe their eggs and chicks, hidden on the ground in tall grass or reeds, were easily nabbed by introduced cats and rats; maybe adults were easily caught in snares. These mechanisms, however, might not as readily affect a bird that nests in tree-tops, or a bird of prey. And for this reason—this selectivity—they provide us a way to link patterns with processes. Selective extinctions, it turns out, can leave traces in the structure of phylogenies that are very different from those of mass extinction events, where species are felled indiscriminately.
To understand why, imagine these rails as leaves of a phylogeny of all bird species across the Pacific (not just on Guam). Then imagine clipping off the tips of the tree one at a time, with shears. At first, as you worked through the fruit doves, honeyeaters, and lories, with unlucky species chosen at random, the tree might retain its overall shape, looking only slightly hollower for your handiwork. But then the shears reach the rails, and you begin pruning more aggressively, lopping off thick branches. The end result would be a tree much more lopsided than the one you started with—a tree with a large chunk taken out of it, a visible hole where there once was a rich assortment of species.
In the language of phylogenetics, this distortion is known as tree balance, and like the frequency of speciation events in a tree, it can be quantified. In other words, the genealogical information in phylogenies can be found not only in the catalogue of kinships but also in comparisons of how extinction has shaped groups of species in different parts of a tree. If the thesis is speciation, its antithesis is extinction. Their synthesis is tree balance.
The usefulness of tree balance in practice is subject to caveats and assumptions, and it deserves substantial caution in interpretation. Tree balance can’t tell you exactly why a given species went extinct; at best, when conditions are right, it can help you guess at how an extinction event occurred. Nor should the signals in tree balance be taken to mean there are fixed differences in competitive ability between different groups of organisms, between doomed rails and conquering snakes; these differences are entirely situational, the outcome of conditionality on conditionality, of the circumstances of particular populations in particular places at particular times.
* * *
Without Charles Darwin, there would be no phylogenies, and though he predated their modern form by a century or so, there’s reason to think he would have appreciated our understanding of how extinction shapes them. “No one can have marvelled more than I have done at the extinction of species,” he boasted in Origin. Extinction, according to Darwin, was a gradual process, one with constraints. If it seemed otherwise, that was the fault of human hubris:
Nevertheless so profound is our ignorance, and so high our presumption, that we marvel when we hear of the extinction of an organic being; and as we do not see the cause, we invoke cataclysms to desolate the world, or invent laws on the duration of the forms of life!
It’s a view of extinction that decentralizes human agency; claims that the process could be quick or catastrophic, Darwin suggests, are a sign of the towering heights of our presumption. The irony is that Darwin’s own theory is an inextricable part of the broader scientific revolution that dramatically expanded human ability to wreak catastrophe on the natural world. This ability was not equally distributed—it was disproportionately concentrated and wielded by the West, and it dovetailed with the expansion of colonialism and genocide. What these forms of violence share is their ability to sunder bonds: bonds between people and places, among family members, between species united by ancestry or ecological relationships.
There is a famous thought experiment in evolutionary biology that asks if whether you replayed the tape of life you would end up with a roughly similar world to our own—with species that resembled Guam’s birds, or brown tree snakes, or ourselves. The exercise is meant to address the question of whether evolution is deterministic, or subject to contingencies, to historical accident. Like any good thought experiment, it can only be addressed by differentially weighting insufficient evidence. The answer folds in on itself; for myself and most other biologists I know, it’s a resounding “it depends.”
You could perform the same thought experiment with extinction, asking whether the rails of some parallel universe are perishing or flourishing. But I don’t think you would be any more successful. In phylogenies of extant species, speciation rates are biased by extinction, and extinction is invisible without speciation. The phenomena are meaningless in isolation, and so their contingencies multiply into a dazzling array of possible outcomes.
Some of these outcomes are already fixed. We cannot resurrect the dead. We can only mourn them, and grapple with the universe we have. The chuguangguang and ko’ko’ met a cataclysm, and their world was desolated. The signature of this cataclysm is written in trees. 